中国癌症杂志
2025, 35 (
):
1067-1075.
10.19401/j.cnki.1007-3639.2025.11.009
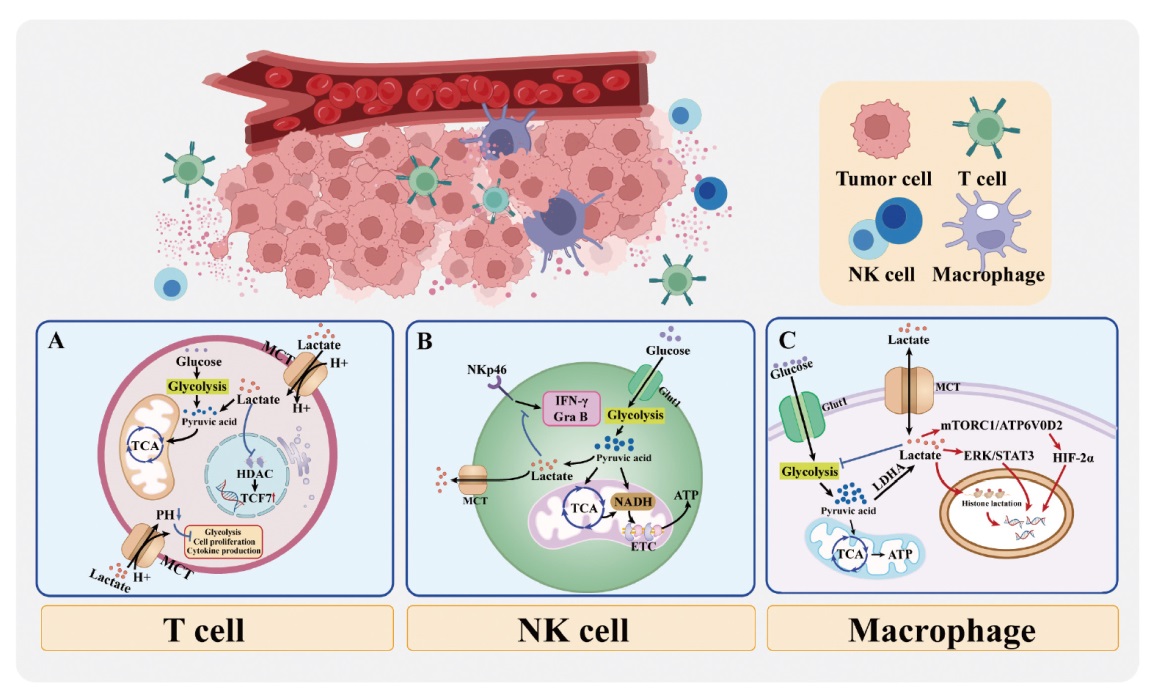
恶性肿瘤作为全球公共卫生领域的重大挑战,迫切需要创新诊疗策略。乳酸作为肿瘤细胞糖酵解的重要代谢产物,不仅是能量代谢中间体,更以“代谢底物”和“信号分子”的双重身份参与肿瘤恶性进程的调控。具体而言,乳酸可通过单羧酸转运体(monocarboxylate transporter,MCT)介导的“乳酸穿梭”实现代谢物跨细胞分配,并借助组蛋白乳酰化修饰调控表观遗传,从而构建能量代谢、氨基酸代谢及脂代谢网络增强肿瘤细胞的代谢可塑性。在免疫调控层面,乳酸诱导T细胞向免疫抑制表型转化,阻碍CD8+ T细胞记忆分化并削弱细胞毒性。同时,乳酸不仅能够降低自然杀伤(natural killer,NK)细胞的免疫效能,还通过诱导线粒体功能障碍触发凋亡,为转移灶创造免疫豁免生态位。此外,高水平乳酸激活多条信号转导通路,募集并诱导巨噬细胞向M2型转化,构建免疫逃逸微环境。治疗策略方面,当前研究围绕乳酸代谢的关键环节展开多维度探索。抑制乳酸合成已成为逆转肿瘤代谢优势的重要治疗策略,通过靶向调控乳酸生成和转运的关键分子,减少肿瘤微环境(tumor microenvironment,TME)中乳酸的蓄积,从而削弱其对代谢重编程的驱动作用。此外,促进乳酸分解是当前极具潜力的代谢干预新方向,借助于生物酶或仿生催化系统,新型药物能够增强局部乳酸的清除效率,缓解酸性TME对免疫功能的抑制。本文系统综述乳酸介导的代谢-免疫调控网络,并探讨靶向乳酸通路的精准治疗策略,旨在打破肿瘤细胞的代谢适应性优势与免疫逃逸之间的恶性循环,为探索更高效的抗肿瘤疗法提供系统性的研究方向和转化思路。
{kind=link}